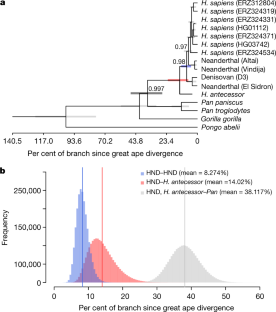
human lineages. (doi:10.1038/s41586-020-2153-8)
This page will be expanded at some point: it is for now only a few salient points giving a global perspective in which to understand the archaeological record of Indian stone age.
Look here for a short account of our evolution from primates, and here for the much larger story of evolution of life itself.
Both from paleontological and genetic evidence we think that the lines separating humans from chimpanzees separated in Africa towards the end of Miocene (23.3–5.2 Ma BP; Ma is short of 1,000,000 years; BP is before present; see here for a short description of the various periods in the history of the earth)—the earliest fossils found to date that are ‘mosaics’ with human and chimpanzee features are from 6–7 Ma BP and molecular estimates date the separation to 5–7 Ma BP. It is likely that the ancestors of the other great apes: gorillas and orang-utangs had branched off earlier than that (10 Ma and 16 Ma respectively). In India, we find Ramapithecus/Sivapithecus (which maybe sexually dimorphic forms of the same species: male Ramapithecus and female Sivapithecus), a probable ancestor of orang-utangs, and some species related to Dryopithecus (these ones probable ancestor of gorilla) from upper miocene (Ramapithecus may have been present as early as 12–14 Ma, the Dryopithecus species may be from around 6 Ma; miocene lasted from 23.3 Ma–5.2 Ma); and Paleosimia, a possible ancestor of orang-utangs, from middle miocene.
The evolution of humans, like those of most species, did not proceed in a linear fashion: rather, random and adaptive changes in Africa, possibly in response to changing environmental and ecological situations, gave rise to a number of species with partly ape-like, partly human, and some novel, characteristics. Most of these have been found east of the rift valley (though some of the earliest specimens are exceptions), and it has been hypothesized that the geographical separation and slow desiccation of this region around 4 Ma BP provided the required impetus. All of these species died out, but since their diversification, earth was probably home to more than one hominid for all but the last thirty thousand years. Naturally, many of these species are unlikely to have been closely related to our direct ancestors, but neither the evolutionary relationships between these groups, nor their taxonomic status is clearly established. Roughly speaking, facultative bipedal large toothed Australopithecus, which may have arisen out of small-toothed ancestors like Sahelanthropus, Orrorin, or Ardipithecus, gave rise to the robust facultative bipedal very large toothed Paranthropus who died out, and the gracile obligate bipedal small toothed Homo which survived.
A word of caution is in order for those unused to thinking at these historical depths: on the scale of human historical processes, speciation is a slow process in which gene flow is slowly quenched by reproductive isolation, either precoital (‘preference’ or ‘behavorial’, ecological, mechanical, or temporal) or postcoital (prezygotic or postzygotic ‘incompatibility’, mortality, reduced fertility, or isolation of the hybrids by any of these means). In organisms with obligate sexual reproduction, establishment of a detectably positive assortative system in experimental conditions takes fifty to a hundred generations in the absence of unusual situations like strong sexual preference or intracellular symbionts; and a rigid reproductive barrier is expected only on time scales at least a thousand times longer. Asian and African subspecies of lions have been separate for some 20–30,000 generations and lions and tigers still produce fertile hybrids after more than 200,000 generations of isolation! Moreover, this need not happen at identical times for all individuals in a species: apart from the vast array of genetic inheritence patterns that can appear, one can also have curious geographic structures like ring species, or situations with hybrid infertility affecting only one of the sexes. There is some evidence that the separation of human and chimpanzee lineages might have started allopatrically, but the complete speciation probably took about 4 Ma (8–4 Ma BP) proceeding through a stage of male hybrid infertility. Given this, the line back through Sahelanthropus, Orrorin, and Ardipithecus described above may not have been the sole contributors to our genetic heritage, and these ancestors, in turn, may have contributed somewhat to the Chimpanzee lineage.
With these caveats, the following table shows a rough guide to the fossil species accepted today (July 2002), though caution is advised since little is known of the ancestors of small toothed Gorilla and Pan (Chimpanzee) that must have been present at the same time!
| Species | Rough Date Ma BP | Species | Rough Date Ma BP |
|---|---|---|---|
| S. tchadensis | 6–7 | K. rudolfensis | 2–2.5 |
| O. tugenensis | 5.7–6.1 | H. habilis | 2–2.5 |
| Ar. ramidus kadabba | 5.2–5.8 | P. robustus | 1.2–2 |
| Ar. ramidus ramidus | 4.4 | P. boisei | 1.2–2.3 |
| A. anamensis | 3.9–4.2 | H. ergaster | 1.6–1.9 |
| A. bahrelghazali | 3.5 | H. erectus | 0.05–1.6 |
| K. platyops | 3.2–3.5 | H. antecessor | 0.8 |
| A. afarensis | 3–4 | H. heidelbergensis | 0.8 |
| A. africanus | 2.5–3.3 | H. neanderthalensis | 0.03–0.2 |
| P. aethiopicus | 2.5–2.7 | H. sapiens | 0–0.2 |
| A. garhi | 2.7 |
Homo could wield stone and bone tools which have been left behind as artifacts, and late in Pliocene (5.2 Ma–1.64 Ma), early man (Homo erectus, probably an offshoot of Homo ergaster) dispersed to different parts of the world (earliest evidence dates to 1.75 Ma BP, surprisingly primitive and small bodied), bearing stone and bone tools. Though these people created the paleolithic tools all over the world, neither they nor their successors the ‘archaic Homo sapiens’ who seem to appear in various parts of the world after about 500–300 Ka BP (Ka is short of 1,000 years) seem to have left any major genetic imprint on modern humans (Homo sapiens sapiens), who might be a separate line from Homo ergaster via Homo heidelbergensis. Recently a study claims to find some genetic evidence in modern humans going back to a population that spread from Africa around 500 Ka BP, and, probably, even earlier; so there might have been limited interbreeding of these people with the later wave described next. But the major genetic component, the ‘last common ancestor of modern humans’ probably lived in Africa around 300 Ka BP; less than 100,000 of them probably came out of Africa to the middle east about 100 Ka BP and started spreading around the world before 65 Ka BP; and there is some indication that what made us truly human was a genetic change that allowed for adult symbolic thought and that arose sometime not much before 50 Ka BP. Isolated modern facial features like the Zhirendong mandible do appear in Asia before this period, but are possibly examples of mosaic evolution of hominins.
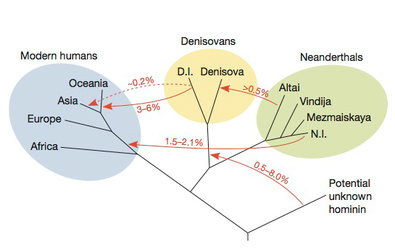
A study reported on in the New York Times in
December 2013 estimated percentages
of genetic transfer between
human species. The split between modern humans and
the
Denisovan/Neanderthals is estimated to have been about 600 kiloyears
back, and
a study from
2012 had found the date of Neanderthal admixture
in humans was
probably between 47–65 kiloyears ago.
The extreme form of this currently predominant view is called the ‘out of Africa’ view, whereas the alternative of a very significant contribution to the modern gene pool from the earlier waves of African emigrants is called the ‘multiregional’ view. Little evidence exists for the latter except some poorly attested possibility of paleontological continuity in certain skull traits, and a disputed example of hybrid neanderthal/sapiens characteristics. Some limited amount of cross-breeding between the ‘modern’ and ‘archaic’ humans however has always been consistent with the evidence (depending on one's bias, this is often called the ‘soft multiregional’ or ‘soft out-of-Africa’ position), though a mitochondrial study favours a gene flow of less than 0.001% per generation in Europe. It had been difficult to actually quantify the ‘limited’ amount or to come to a definite conclusion without a large scale genomic study of multiple unlinked genes, and without understanding the selective sweeps and the population history. In fact previous estimates of about 15% contribution of older DNA into non-Africans now seem exaggerated on the strength of direct Neanderthal sequences: The best estimates now is that may be 1–4% of the genome of non-Aricans derives from early admixture from Neanderthals. Similarly, Denisovans, who are a splinter group from the line leading to Neanderthals, seem to have contributed 4–6% to Melanesians, perhaps explaining some of the Australoid facial features of these Archaic humans remarked on previously. Recent work (see figure) is showing a complicated interchange of genetic material between archaic human species. Nevertheless the complicated distribution of the admixture suggests that the interbreeding was not easy: male children of Neanderthal mothers and modern human fathers were possibly infertile. And even though some Neanderthal traits related to immune system and skin structure may have been selected for in modern humans, others gave rise to diseases like Crohn's disease and Type II diabetes.
All this is qualitatively consistent with earlier coalescense studies on isolated loci, but those studied were limited by large statistical errors, and results were, therefore, difficult to interpret. I describe here, one such study that may indicate the general pattern. This study has claimed that some genes (CYP1A2, FUT2, Lactase) might trace back to the homo erectus expansion out of Africa 1.9 (0.99–3.10) Ma BP. Some other genes (CCR5, EDN, FUT2, FUT6, PDHA1) may have spread due to cross-breeding amongst the different geographical populations over the next 1.25 Ma. Some 650 (390–970) Ka BP, there may have been a separate Acheulian expansion out of Africa (the Neanderthal expansion seen in genes like FUT6, G6PD, Haemoglobin β, HFE, Lactase, MS205, MC1R), and a number of genes (CYP1A2, ECP, G6PD, HFE, Haemoglobin β, MSN/ALAS2, RRM2P4, Xq13.3) may show a diffusion over the next 500 Ka. The major ‘out-of-Africa’ took place around 130 (96–169) Ka BP and is evident in the mtDNA and Y chromosomes, as well as some other genes (HFE, HS571B2, RRMP24). At a later date, there may even have been a back migration from South Asia to Africa which affected some genes like Haemoglobin β and affected Y chromosome data. The rest of the story is the story of modern human movements from South Europe to North Europe, from South Asia to North Asia to the Americas, from South Asia to the Pacific (shown in genes like EDN, MS205, MC1R, MX1, TNFS5F as well as mtDNA data), as well as gene flow amongs these modern populations which we see all over the genome. More recently, there were studies which had identified features like the H2 clade at the MAPT locus, or the D haplotype of the encephalin gene as belonging to the Neanderthal lineage; or variants of RRM2P4 in Chinese as indicative of homo erectus population admixtures; but the explicit sequencing of the Neanderthal genome has not confirmed these.
![]()
![]()